During photosynthesis, plants incorporate atmospheric CO2 into carbohydrates using the energy of sunlight. CO2 enters the leaf through pore-like openings located predominately in the lower epidermis of the leaf called stomata. The sizes of the stomata are directly controlled through regulating the turgor pressure of the surrounding cells, known as guard cells. By inflating and deflating the guard cells, the plant can optimize the balance between CO2 uptake and water loss in real time as environmental conditions change.
Stomatal Opening
Stomata open when the pair of guard cells is fully turgid and pressing firmly against each other. Although this seems counterintuitive, the pressurized condition results in open stomata because of the unique anatomy of these specialized cells. In particular, the interior walls of the guard cells are heavily reinforced with cellulose, making them bow open when stressed by the influx of water. But what causes this influx of water? As usual, water flows in response to the water potential gradient.
In sunny conditions, blue light stimulates a H+-ATPase in the guard cell plasma membrane, which hydrolyzes ATP and pumps H+ out of the cell. The outward flow of H+ results in the inside of the cell becoming more and more negatively charged. This hyperpolarization leads to the activation of an inward-rectifying K+ channel, resulting in the influx of K+ ions. This influx of K+ activates a H+/Cl- symporter, which imports Cl- ions. The net uptake of both K+ and Cl- significantly reduces the water potential, resulting in the influx of water from the apoplast.
Stomatal Closing
Stomata close in response to a number of different signals. One such signal is water stress, usually signaled through the presence of the drought hormone, abscisic acid (ABA). The perception of ABA in the guard cell leads to the release of intracellular Ca++, which triggers the opening of a plasma membrane anion channel. This allows the efflux of Cl- ions, leading to the depolarization of the membrane. Upon membrane depolarization, an outward-rectifying K+ channel opens and allows the escape of K+. The loss of both K+ and Cl- increases the water potential of the guard cell, leading to water efflux.
One important question is, where are the large supplies of ions and water coming from that lead to stomatal opening, and where do these go when stomata close? Guard cells do not exist in isolation, but are found situated in the epidermis next to companion cells. As the concentration of ions is increasing in guard cells, those ions are being supplied by companion cells. Likewise, the ions that flow out of guard cells undergoing stomatal closing are taken up by the companion cells.
The major export product from photosynthesis is glyceraldehyde-3-phosphate (G3P), a triose phosphate carbohydrate, which can enter either the starch or sucrose biosynthesis pathway depending on conditions in the cell. During the daytime, much of the carbon that is fixed by photosynthesis remains in the chloroplast and enters the starch biosynthesis pathway. At night, carbon stored in the form of starch is mobilized by conversion to sucrose, which is synthesized in the cytoplasm.
Starch Biosynthesis
Starch, formally known as α-amylose, is a long-chain polysaccharide made of α 1→4 linked glucose, where the chain length numbers in the hundreds or thousands. α-amylose forms a single helix structure because of its regular repeating pattern, and this secondary structure readily crystallizes. The first step in the synthesis of α-amylose is the formation of hexose phosphates, including fructose 6-phosphate, glucose 6-phosphate, and glucose 1-phosphate. Glucose 1-phosphate is further ‘activated’ by reacting with the sugar nucleoside ATP to produce ADP-glucose. This form of glucose is highly reactive and readily joins an elongating chain of α-amylose at the 4-carbon position to give the characteristic α 1→4 linkage.
While α-amylose represents about 30% of the total starch in most plants, the rest of the starch is in a highly branched form called amylopectin. Rather than forming straight chain helices that readily crystallize, amylopectin does not crystallize. Amylopectins are formed by starch branching enzymes that form branches among short α-amylose chains that are α 1→6 glycosidic bonds.
Sucrose Biosynthesis
When triose phosphates are exported from the chloroplast, they enter the sucrose biosynthetic pathway in a similar manner as the start of the starch pathway — by condensation to form a pool of hexose phosphates. Also like starch biosynthesis, glucose 1-phosphate reacts with a sugar nucleoside, in this case UTP instead of ATP, to form UDP-glucose. Sucrose is the result of the condensation reaction between this UDP-glucose and fructose 6-phosphate. This sucrose serves as the major form of transportable carbohydrate within the plant.
Regulation
What determines whether the triose phosphates formed by photosynthesis enter the starch or sucrose pathways is the activity of a chloroplast envelope transporter. This transporter, called the triose phosphate-Pi antiporter, exchanges triose phosphates for Pi (inorganic phosphate) between the stroma and cytoplasm. When concentrations of Pi are high in the cyctoplasm, the antiporter is activated and exports triose phosphates in exchange for the uptake of Pi, with the cytoplasmic triose phosphate entering sucrose synthesis. On the other hand, when cytoplasmic Pi is low, no exchange happens and triose phosphates remain in the chloroplast to enter the starch synthesis pathway.
Despite some apparent benefits of photorespiration, several taxa of plants have evolved strategies that minimize photorespiration. These additional pathways for carbon fixation evolved in plants growing under high light conditions with limited water resources and took advantage of proteins from other primary metabolic processes, co-opting them for a new purpose. These pathways exist in addition to the process of photosynthetic carbon reduction already described, serving to concentrate CO2 for incorporation by Rubisco rather than operating instead of Rubsico.
C4 Pathway
One additional pathway that concentrates CO2 for Rubisco is known as the C4 pathway, and it is found in some members of the grass (Poaceae) family such as maize, sorghum, and sugar cane. These grass species display a specialized anatomical adaptation consisting of a ring of cells surrounding leaf vascular bundles called the bundle-sheath cells. The expression of Rubisco and other genes encoding enzymes of the Calvin-Benson pathway are limited to the bundle-sheath cells, resulting in the spatial separation of initial carbon fixation and Rubisco activity. The initial fixation of CO2 happens in the mesophyll cells of the leaf, and is carried out by an enzyme usually associated with glycolysis called phosphoenolpyruvate (PEP) carboxylase. Following carbon fixation, the resulting 4-carbon molecule (malate) is transported into a bundle-sheath cell and decarboxylated, giving off a CO2 that will be incorporated by Rubisco into the Calvin-Benson cycle.
The formation of a gradient never occurs without the input of energy, and this process of concentrating CO2 in the bundle-sheath is no exception. C4 plants invest heavily in the form of ATP and NADPH to sustain the gradient, but because this pathway is found in plants in high light environments, this cost is easily absorbed. Ha.
CAM Pathway
Unlike C4 plants, CAM plants do not have any particular anatomical adaptations related to carbon fixation. CAM plants employ the same alternate enzymes as the C4 plants for initial carbon fixation, but place them under the control of circadian regulation, restricting carbon fixation to the night, when stomata are open. This arrangement allows the plant to exchange gas with the environment when temperatures are much lower, resulting in significant water savings. As CO2 is fixed by PEP carboxylase, the product (malate) is imported to the vacuole and stored as malic acid. During the daytime, when stomata are closed, this malate is exported from the vacuole and decarboxylated, increasing CO2 concentrations and favoring its use over O2 by Rubisco.
Regulation
PEP carboxylase activity is regulated by two main mechanisms. High concentrations of malate cause a form of enzyme inhibition known as feedback inhibition, in which the product of a pathway exerts an influence over an enzyme upstream of its production. Levels of malate are kept low in the vicinity of PEP carboxylase by exporting it to the bundle-sheath cells in the case of C4 pathway plants or importing it into the vacuole in the case of CAM plants. Another means of regulation over these alternative carbon fixation pathways involves the phosphorylation of PEP carboxylase, which occurs in the light. Upon phosphorylation, the affinity of PEP carboxylase for the substrate phosphoenolpyruvate increases. The affinity of an enzyme for its substrate, known as the Michaelis constant (Km), can be empirically determined in a carefully-controlled in vitro experiment. A decreasing Km is indicative of greater substrate binding at a lower substrate concentration. Conversely, the phosphorylation of PEP carboxylase causes an increase in the inhibition constant (Ki) for malate, meaning that the same amount of malate would inhibit the enzyme less under phosphorylated conditions.
The enzyme responsible for incorporating atmospheric CO2 in the reactions of photosynthesis is called Rubisco, an ancient, large, multi-subunit enzyme with a major apparent weakness. In addition to incorporating CO2, Rubisco can also bind O2 and perform an oxygenation reaction with the substrate ribulose bis-phosphate. The results of this oxygenation reaction are one molecule of phopshoglycerate and one molecule of a 2-carbon product called phosphoglycolate. While the PGA can enter the reduction pathway in the chloroplast, the phosphoglycolate cannot and instead enters a recovery pathway spread across two other organelles, the peroxisome and mitochondria. The oxygenation reaction together with the various recovery steps are known as the photorespiratory pathway and represent a significant energetic cost to the plant.
There are probably many explanations for the existence of this costly pathway, beginning with a consideration of the conditions under which Rubisco evolved. Photosynthetic carbon reduction probably began over 1 billion years ago, and is itself thought to have been responsible for the formation of most of the O2 in the atmosphere. In other words, in the earliest cases, there was little to no competition between CO2 and O2 for the active site of Rubisco. So the simplest explanation for why oxygenation happens is that there was not a selective pressure to discriminate between these two substrates. Another explanation is that, given the integration of the recovery pathway with various metabolic pathways such as amino acid biosynthesis, it represents a source of inputs to these pathways. A third explanation is that photorespiration is a strategy for the plant to cope with excess light energy under conditions in which CO2 is limiting. Such conditions arise when the plant is water-stressed and closes stomata to conserve water, which results in a build-up of O2 and draw-down of CO2. Evidence for photorespiration as an important energy-shunting pathway comes from experiments in which mutants for various steps in the recovery pathway were isolated and grown under various conditions. Mutants grow normally under low-light conditions, but when challenged with light levels similar to full sun the mutants show symptoms consistent with damage due to photoinhibition.
Below is a flow diagram of the Calvin cycle, found in the chloroplasts of photosynthetic organisms. Each substrate and enzyme is clickable, and leads to a site with detailed information on the participant. A PDF of this flowchart is also available for printing.
The carbon reactions of photosynthesis, also known as the Calvin-Benson cycle or photosynthetic carbon reduction, consist of a series of steps that take place in the chloroplast. Broadly speaking, the carbon reactions consist of three steps: carbon fixation, reduction, and regeneration of the initial substrate of the cycle. Because the carbon reactions together form a metabolic cycle, there is no end product of the pathway as such. Rather, the cycle ‘exports’ reduced carbon in the form of triose phosphates.
Carboxylation
In the first step of the carbon reactions, CO2 reacts with a high-energy five carbon substrate called ribulose bisphosphate (RuBP), resulting the the formation of two molecules of the three carbon compound phosphoglycerate (PGA). This carboxylation reaction is catalyzed by Rubisco, a large and evolutionarily ancient enzyme made up of 8 large and 8 small subunits. Rubisco is one of the most abundant proteins on Earth, and makes up 40% of the soluble protein in an average leaf. Rather than attaching the CO2 onto the end of the five carbon substrate, Rubisco adds it to the second carbon in the chain, forming a very unstable intermediate that splits between the 2 and 3 carbons almost immediately. The resulting PGA molecules enter the reduction phase of the cycle.
Reduction
The first step of the reduction phase of the carbon reactions is where the majority of the energetic compounds from the light reactions are used. In the first reaction of reduction, PGA is phosphorylated to make bisphosphoglycerate, a reaction that uses ATP produced by the light reactions. In the second reaction of the reduction phase, bis-phosphoglycerate is reduced to glyceraldehyde-3-phosphate (G3P) by the donation of electrons from NADPH, also a product of the light reactions. G3P is a triose phosphate carbohydrate and is the true product of the carbon reactions in the sense that it is exported from the cycle and enters the starch or sucrose biosynthetic pathways.
Regeneration
The rest of the carbon reactions, 10 individual enzyme-mediated reactions, make up the regeneration phase of the cycle. The complexity of the regeneration phase is due to the difficult task it is given: to make a highly reactive five-carbon molecule starting with a three-carbon molecule. To accomplish this, the regeneration phase employs a network of interrelated reactions in which the pool of triose phosphates serve as reactants many different times.
Regulation
The carbon reactions are regulated by two key factors: the pH of the stroma and a redox regulatory feedback loop. Although the carbon reactions are sometimes referred to as the ‘dark reactions’ to illustrate their separation from the light reactions, this is a misnomer. The carbon reactions are highly dependent on light, not just for the products of the light reactions that act as substrates, but also for regulation of several key enzymes. Three enzymes in particular have a pH optimum around 8, including Rubisco (the other two are fructose 1,6-bisphosphatase and sedoheptulose bisphosphatase). This high pH requires the transport of protons out of the stroma and into the lumen, thus the maximum activity of these enzymes occurs in the light.
A second point of regulation of the carbon reactions is a redox regulatory feedback loop. If you recall, PS I reduces an iron-sulfur protein called ferredoxin (Fd), which can supply electrons to NADP+. In addition, it can reduce another small redox-active protein called thioredoxin. When reduced, it can reduce disulfide bridges that exist in several enzymes in the carbon reactions, changing them from an inactive state to an active state.
Efficiency
To calculate the overall efficiency of photosynthesis, we need to correlate the input energy from light with the output energy in the form of carbohydrates. On average, it takes 8 photons of light to fix one molecule of CO2 via the reduction of 2 NADP+ molecules and the synthesis of 3 ATPs. Thus it will require an input of 48 photons to fix one molecule of a hexose carbohydrate (six CO2 fixation events). If we assume each of these photons contains the minimum energy necessary to drive photosynthesis, about 680 nm light, that would be 175 kJ per mol of photons. Therefore, multiplying 48 mol × 175 kJ per mol gives 8400 kJ of energy for 1 mole of hexose carbohydrate. This represents the minimum energy required by the plant to produce a mole of hexose. When fully oxidized by the cell, this carbohydrate yields 2804 kJ of energy, giving an overall theoretical efficiency of 33% (2804 / 8400).
The photosynthetic light reactions take place across the thylakoid membrane, an extensive system of membranes extending throughout the chloroplast. This network of membranes is home to a variety of membrane-spanning proteins involved in the interception of light or the transfer of electrons. Working together, these proteins transduce the energy of light into an electrochemical gradient across the thylakoid that ultimately drives ATP synthesis. At the same time, they form a series circuit that removes electrons from water and donates them to an electron acceptor called NADP+, producing NADPH.
Light Harvesting
While there is a variety of pigment molecules involved in photosynthesis, light harvesting and energy transduction are carried out by the chlorophylls. Chlorophylls are closed-ring tetrapyrroles that coordinate a Mg+ in the center of the ring that plays a role in the redox reactions at the core of photosynthesis. In addition, they have a long fatty-acid tail that renders them hydrophobic. Chlorophylls interact with light and show two peaks in their absorption spectra: one in the red range and another in the blue.
Chlorophylls are not freely distributed in the thylakoid membrane, but are found in association with polypeptides. One such class of protein is known as the Light Harvesting Complex, or LHC proteins. These proteins act as antenna complexes, gathering light energy and directing it to the reaction center chlorophylls. In other words, the chlorophylls associated with the antenna complex proteins are not themselves chemically active — that role is reserved for the reaction center chlorophylls. On average, only 1 in 250 chlorophyll molecules is chemically active.
The chlorophylls associated with antenna complex proteins are nonetheless critical to photosynthesis, because as they are struck by photons and excited, they are capable of transmitting that excitation to nearby chlorophylls through resonance energy transfer. Research has shown that this process of energy transfer is an example of quantum coherence, and the network of chlorophylls acts in concert to transfer excitation energy with near 100% efficiency to the reaction center chlorophylls.
Reaction Centers
The LHC proteins surround photosynthetic reaction center proteins. The reaction center proteins also have a number of chlorophylls associated with them that receive the excitation energy from LHC chlorophylls. In addition, each of the 2 reaction center complexes has a set of chlorophylls that are chemically active — they are capable of photochemistry. These chlorophylls, known as the reaction center chlorophylls or special pair, transfer an electron to an electron acceptor upon excitation. When in the ground state, prior to absorbing light energy, the reaction center chlorophyll special pair is both a poor reducing agent and a poor oxidizer. But when excited by energy absorbance, it becomes both a good reducing agent and oxidizer.
Evidence for two photosynthetic reaction centers in plants came from a series of experiments carried out by Robert Emerson. Working with the green alga Chlorella, Emerson used flashes of light at specific wavelengths to drive photosynthesis while measuring the net yield of O2. He observed a particular rate of O2 yield under both 670 nm and 700 nm light. When the culture of algae was excited by both wavelengths simultaneously, however, Emerson measured a rate of O2 yield that was greater than the sum of O2 produced by either wavelength alone. This simple experiment had profound implications, as Emerson and many others would go on to show that photosynthetic electron transport occurred through a series of two reaction centers in series. In other words, electrons passed from one of the reaction centers to the other through a number of intermediaries in and around the thylakoid. The two photosynthetic reaction centers are known as Photosystem II (PS II) and Photosystem I (PS I).
Photochemistry
The excitation of the reaction center chlorophyll special pair in PS II raises the energy level of this molecular complex to the point that it becomes energetically favorable for it to lose an electron. As a result, the excited-state chlorophyll reduces a nearby molecule of pheophytin, which is nearly identical in structure to chlorophyll save for the Mg+ at its center. This structural similarity ‘tunes’ the pheophytin to be an efficient electron acceptor from the excited state special pair. The positioning of the primary acceptor pheophytin is important as well, as it is found in the reaction center protein toward the stromal face of the membrane spanning domain. This position results in the charge separation across the thylakoid membrane that underlies the production of an electrochemical gradient.
Midpoint potentials of the various redox components in photosynthetic electron transport.
The loss of an electron from the reaction center special pair is known as primary photochemistry, and this leaves the special pair in an unstable, oxidized state. In PS II, the special pair can be re-reduced by a nearby tyrosine amino acid, leaving the tyrosine oxidized. If we follow the trail of redox reactions back to their origin, we find that ultimately, the electrons that re-reduce the special pair are removed from water at a polypeptide closely associated with the lumenal face of the PSII reaction center called the Oxygen Evolving Complex (OEC).
The OEC catalyzes the oxidation of water through a stepwise series of redox intermediate states made possible through its coordination of Mn2+ ions. The OEC thus provides electrons one-at-a-time to the reaction center special pair as they are removed from a cluster of Mn2+ ions. These electrons are eventually replaced on Mn2+ when two water molecules are simultaneously oxidized, releasing four protons (H+) and molecular oxygen (O2). In other words, the oxidizing potential of the reaction center special pair is transferred to the OEC, where it is used to oxidize water.
In PS I, the mechanics of photochemistry occur the same as on PS II, but the primary electron acceptor is a modified chlorophyll rather than pheophytin. Rather than having an OEC as a source of electrons, the electron donor for PS I is a small molecule called plastocyanin, which diffuses to the lumenal face of PS I carrying electrons that ultimately came from water via PS II.
Electron Transport
After the oxidation of the PS II reaction center, the lost electron enters a pathway of electron flow through several complexes embedded in the thylakoid membrane. This electron transport pathway includes the protein complexes PS II, cytochrome b6f, and PS I. While it is convenient to represent each of these three components nearby each other, they are actually localized in different parts of the thylakoid membrane. PS II and cytochrome b6f are in the interior portions of thylakoid stack, while PS I is found in regions of the membrane having access to the stroma. Two pools of electron carriers connect each of the three major components, with a small, hydrophobic organic molecule called plastoquinone diffusing between PS II and cytochrome b6f, while a copper-containing protein called plastocyanin links cytochrome b6f with PS I. In other words, PS II reduces plastoquinone, and plastocyanin serves as the electron donor for PS I.
The three major components of the electron transport system do no exist to simply pass electrons along to NADP. Their orientation within the thylakoid is such that, as electrons travel through the various components, their flow enhances the concentration of protons that forms in the lumen due to water oxidation. As described above, the direction of electron movement during photochemistry on PS II is from the lumenal face toward the stromal face of the thylakoid due to the position of the primary electron acceptor. This directional flow allows plastoquinone to gather two protons from the stroma along with the two electrons it receives from PS II. When reduced, plastoquinone leaves PS II and diffuses through the membrane and binds to a cytochrome b6f complex on the lumenal face. As plastoquinone reduces the cytochrome b6f complex, the two protons it was carrying are released into the lumen.
Cytochrome b6f engages in two different processes with the electrons it receives from plastoquinone: non-cyclic transport and cyclic electron transport. When performing non-cyclic transport, the cytochrome b6f complex is reduced by plastoquinone and reduces plastocyanin at the lumenal face. Under certain conditions though, cytochrome b6f passes electrons among several intrinsic electron acceptor sites, each of which can coordinate a quinone molecule. Because this process relies on quinone sites, this has been named the Q cycle. The various quinone binding sites on the cytochrome b6f complex facilitate the flow of electrons toward the stromal face, where they eventually reduce a plastoquinone associated with the cytochrome b6f complex. This plastoquinone is identical to that described above, and once reduced it leaves its binding site at the stromal face of the cytochrome b6f complex and binds at the complex’s lumenal face. The end result of the Q cycle is to further enhance the flow of protons into the lumen.
Once reduced by cytochrome b6f, plastocyanin (a single electron carrier) leaves the complex and diffuses through the lumen to PS I. As the electron donor to PS I, plastocyanin is oxidized by the PS I reaction center special pair when it absorbs a photon of light energy and undergoes photochemistry. The electron that is lost from the reaction center travels to a modified chlorophyll and several intermediate electron acceptors within PS I before reducing ferredoxin, bound to the stromal side of PS I. Ferredoxin is a small protein that coordinates an iron-sulfur complex. When reduced, it provides the reducing power to the enzyme involved in reducing NADP+, called ferredoxin-NADP reductase. It can also participate in cyclic flow of electrons between PS I and the cytochrome b6f complex under certain conditions. The net result of the electron transport pathway is the conservation of light energy in the form of an electrochemical potential gradient across the thylakoid membrane, and the reduction of NADP+ to NADPH.
ATP Synthesis
The energy that is represented by the proton gradient across the thylakoid membrane can be used by ATP synthase to catalyze the addition of a phosphate onto ADP to make ATP. The ATP synthase enzyme is a multimeric molecular machine that converts the gradient of protons into kinetic, rotational energy by permitting protons to flow through its membrane-spanning pore. As protons leave the region of high concentration in the lumen and pass through the pore region, they exit into the stroma, which causes rotation of the membrane-spanning domain. This rotation is propagated by a shaft-like domain called the γ domain to the catalytic domain. Within the catalytic domain, ADP and P(i) are bound, and the rotation of the γ domain causes a conformational change that forces the ADP and P(i) to react and produce ATP.
What we call ‘photosynthesis’ is a collection of events that uses the energy contained in light and reducing CO2. A simplified net reaction is shown below:
H2O + CO2 + light → (CH2O)n + O2
On an annual basis, this collection of processes is responsible for fixing 7 × 1013 kg of CO2. To help put this value in perspective, that is equal to 1% of all known fossil fuel reserves on the planet and roughly 10× the annual global energy consumption. In addition, photosynthesis also releases oxygen at a rate that replaces all O2 in the atmosphere every 2000 years.
Photosynthesis consists of 2 interdependent processes known as the light reactions and the carbon reactions. For our purposes, we will further disassemble the light reactions into the processes of photochemistry and electron transport. Both of these parts of the light reactions take place across the system of membranes found within chloroplasts, known as the thylakoid membrane system.
As we will discuss in more detail elsewhere, light results in the polarization of this membrane due to the formation of a H+ gradient. This proton gradient is used to by ATP synthase to combine ADP and Pi to form ATP, which is one of the key products of the light reactions. The other major product of the light reactions is an energy carrier called NADPH. All together, the light reactions can be thought of as a series of oxidation-reduction reactions powered by light energy, with electrons supplied by H2O and ultimately reducing NADP+ to NADPH.
In the carbon reactions, the energy from the light reactions, represented by ATP and NADPH, is used to reduce atmospheric CO2 to carbohydrates. This takes place in a series of enzyme-mediated reactions that takes place in the stroma of the chloroplast. The series of reactions represents an example of a biochemical cycle, with the initial reactant regenerated by the pathway. The products of the cycle are excess molecules that are “exported” from the pathway.
It is fairly obvious from the above descriptions that the carbon reactions are dependent upon the light reactions for ATP and NADPH. But it is important to keep in mind that the ADP and NADP+ produced by the carbon reactions are absolutely essential for the light reactions to continue. Without the return of these substrates to the light reactions, the cell would experience significant damage due to continued exposure to light energy.
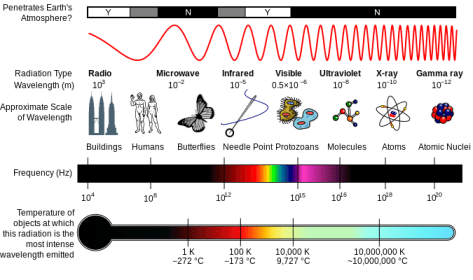
Light is one form of electromagnetic radiation. As such, it occupies a small slice of the electromagnetic spectrum, which also includes forms of radiation having greater energy than visible light, such as gamma rays and x-rays, and other forms having less energy than light, such as infrared radiation and microwaves. Each of these is characterized by two physical parameters that describe the quality of radiation: wavelength (λ) and frequency (ν). Visible light includes radiation having a wavelength between approximately 400 nm and 700 nm. Because the speed of radiation, c, is a universal constant, wavelength and frequency always vary inversely with each other and their product is equal to c.
A diagram of the Milton spectrum, showing the type, wavelength (with examples), frequency, the black body emission temperature.
Because light is a form of energy, and because we are interested in understanding how light energy is transduced into chemical energy, it is important to understand the relation between light quality and energy. Comparing blue light (450nm < λ < 475nm) with red light (620nm < λ < 730nm) for example, note that blue light has a smaller wavelength and therefore a greater frequency than red light. Even though the speed of light is a constant, the energy is not — energy varies in direct relation to frequency. So in the above example, blue light contains greater energy than red light of a given fluence rate.
In addition to being described in terms of its wave-like properties, the energy in light exists in discrete packets (called photons) rather than as a continuous stream. Photons are measurable down to the individual one, known as a quantum. As we will see next, the fact that light travels in photons, each having a predictable quantum of energy, helps us understand the events surrounding light absorbance by molecules and the transduction of energy into a chemical form.
Light Interacts with Molecules
As you look around the world, you see objects of all different colors. The production of those colors, and in fact our ability to perceive them, both rest on the fact that visible light can interact with matter in specific, predictable ways. Whether a particular frequency of radiation interacts with a particular molecule depends on the structure of the molecule — the arrangement of atoms in space. In the same way an aerial antenna depends on the precise spacing of its elements to intercept radio waves (another form of electromagnetic radiation) of specific frequencies, so too are biological molecules ‘tuned’ to specific frequencies of light to maximize energy transduction. In the case of the photosynthetic pigments known as chlorophylls, their molecular structure allows them to absorb blue and red wavelengths, but not yellow, green, or orange. While most plants have pigments that absorb yellow and orange, green light goes unused by the plant, being either transmitted or reflected.
When a pigment molecule intercepts a photon for which it is receptive, the energy of the photon is conserved by a rearrangement of electrons in the molecule. This rearrangement results in the boosting of an electron from the ground state to an excited state. Sometimes, the excited state is dissipated through the re-emission of light energy, known as fluorescence. Other times, if there are like molecules in close proximity, the excited state can be passed among them through resonant energy transfer. Under certain highly specialized circumstances, the excited state can represent the first step toward a photochemical reaction in which the electron departs the excited state molecule and reduces a nearby electron acceptor. Finally, the excited state may be dissipated through a process called radiationless decay, which releases heat to the surroundings. Each one of these processes plays an important role in photosynthesis.