Although the most primitive land plants possessed little more than stem-like structures, most land plants today produce broad, flat leaves that maximize the surface area exposed to light. While it may be clear how meristem-driven growth can result in the formation of an axial organ, like a stem or root, it is probably harder to imagine how a meristem could produce a laminar structure like a leaf.
The first visible indication that a leaf is about to form is a tiny bump on the surface of the apical dome, the tip-most section of the elongating stem. These bumps form off to one side of the dome, and the position of the bump determines the pattern of leaf placement along the stem, called phyllotaxy. In other words, for a species having alternate phyllotaxy, the position of consecutive bumps would spiral around the apical dome, while species with opposite phyllotaxy would form two bumps simultaneously and offset by 180° from each other. As time passes, the bumps, known as leaf primordia, expand into leaves, and the leaves become separated by expansion of the stem over the time between which the bumps formed.
Given that the arrangement of leaves on a stem is highly reproducible within a species, the placement of each leaf primordium must be regulated. Over the last 20 years, there has been substantial progress made in identifying some of the factors that control maintenance of the meristematic population as well as initiation of new leaf primordia. One of the key concepts to emerge is that, while the meristem looks like an amorphous mass of cells, it is actually organized into two zones based on the expression patterns of particular classes of genes, many of which encode transcription factors. These two zones, known as the central zone (CZ) and peripheral zone (PZ), exist in a dynamic tension, such that they tend to protect their cellular ‘turf’ by preventing expression of genes from the other zone. In addition to zones, the shoot apical meristem is also partitioned into layers, with most species having 3 layers (L1, L2, and L3).
Leaf primordia form in the peripheral zone, and even before a bump is visible, the region where a primordium will form shows downregulation of a transcription factor associated with meristem activity, the STM gene in Arabidopsis. Shortly thereafter, this region begins to protrude from the surface as a leaf primordium. The L1 layer, which is the outermost layer of cells, will form the dermal tissue of the new leaf, while the L3 layer will form the vascular tissues. L2 goes on to differentiate into the rest of the tissues that make up the leaf, most of which are ground tissue. As the leaf primordium is beginning to emerge, cell divisions in L1 and L2 occur perpendicular to the surface of the apex, while those in L3 occur in both perpendicular and parallel planes.
In addition to an important role for transcription factors and signal peptides, plant hormones also have prominent roles in the emergence of leaf primordia. Using an auxin-responsive reporter gene, it has been determined that leaf primordia are local sites of auxin maxima. Auxin, or indole-acetic acid (IAA) is synthesized in existing primordia and transported across the apical dome, where it appears to accumulate and coincide with the site of primordium formation. This accumulation requires a system of polar auxin transporters, which have now been shown to direct auxin transport across the shoot apex to the site of incipient primordium formation. As the new primordium emerges, it exports the auxin necessary to form a new maximum on the other side of the meristem, where a new auxin maximum will form. In other words, as primordia mature, they become auxin sources rather than auxin sinks.
Reference
Barton MK (2010) Twenty years on: The inner workings of the shoot apical meristem, a developmental dynamo. Developmental Biology 341:95-113
 = 0)
= 0) ) represents the effect of any solutes on the free energy of water. The addition of a solute to water decreases the concentration of water, thus reducing its free energy. The degree of this reduction is a function of the final concentration of solute particles:
) represents the effect of any solutes on the free energy of water. The addition of a solute to water decreases the concentration of water, thus reducing its free energy. The degree of this reduction is a function of the final concentration of solute particles: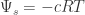
 ) represents the effect of pressure on the free energy of water; positive pressure increases free energy, and negative pressure reduces it. Pressure within a typical leaf or root cell is positive, while that within the xylem is negative. A cell with
) represents the effect of pressure on the free energy of water; positive pressure increases free energy, and negative pressure reduces it. Pressure within a typical leaf or root cell is positive, while that within the xylem is negative. A cell with 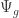 ) in such a plant. Each 10 m in vertical distance translates to a 0.1 MPa change in water potential.
) in such a plant. Each 10 m in vertical distance translates to a 0.1 MPa change in water potential. ) plays a significant role in determining the flow of water. When water is bound to a surface in a very thin layer, that water is less available to do work, reducing its free energy. This parameter plays an important role during seed germination and under drought conditions.
) plays a significant role in determining the flow of water. When water is bound to a surface in a very thin layer, that water is less available to do work, reducing its free energy. This parameter plays an important role during seed germination and under drought conditions.

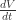 , is equal to the product of effective pressure (turgor pressure (P) minus wall yield threshold (Y)) and wall extensibility (m). This is a version of the Lockhart equation, first published in 1965
, is equal to the product of effective pressure (turgor pressure (P) minus wall yield threshold (Y)) and wall extensibility (m). This is a version of the Lockhart equation, first published in 1965


